|
 |
 |
|
|
|
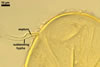
SPORES occur singly in the soil; origin blastically at the tip of hyphae continuous with extraradical mycorrhizal hyphae. Spores yellowish white (4A2) to butter yellow (4A5); globose to subglobose; (92-)108(-146) µm diam; or ovoid; 104-115 x 123-139 µm; with one subtending hypha.
SUBCELLULAR STRUCTURE OF SPORES consists of a spore wall composed of two tightly adherent layers (swl1 and 2).
 |
 |
 |
 |
 |
 |
 |
 |
 |
In PVLG |
In PVLG+Melzer's reagent |
|||||||
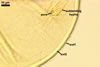
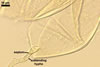
Layer 1, forming the spore surface, semi-permanent, semi-flexible, hyaline, (0.7-)1.0(-1.2) µm thick, frequently intact or only slightly deteriorated in mature spores, sometimes associated with granular soil debris.
Layer 2 laminate, semi-flexible, smooth, yellowish white (4A2) to butter yellow (4A5), (2.5-)3.5(-4.4) µm thick.
None of the two layers stains in Melzer's reagent.
 |
 |
 |
 |
 |
 |
In PVLG |
|||||
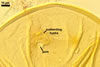
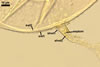
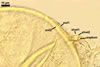
Wall of subtending hypha yellowish white (4A2) to butter yellow (4A5); (1.0-)1.6(-2.7) µm thick at the spore base; composed of two layers continuous with spore wall layers 1 and 2.
Pore 3.8-5.1 µm diam, open or occluded by a straight or recurved septum, (1.0-)1.4(-2.0) µm thick, continuous with the innermost laminae of the laminate spore wall layer 2, positioned up to 11.3 µm below the spore base.
GERMINATION. Not observed.
MYCORRHIZAE. The presence of mycorrhizae in field-collected root samples was not determined.
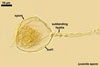
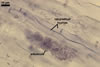
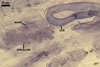
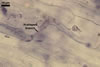
In one-species cultures with Plantago lanceolata L. as the host plant, mycorrhizae of Gl. eburneum comprised arbuscules, as well as intra- and extraradical hyphae. Arbuscules consisted of short trunks branched from parent hyphae and numerous branches with very fine tips. Their distribution along root fragments ranged from uniform to patchy, depending on the root fragment examined. Intraradical hyphae grew along root axis, were (2.0-)4.9(-8.3) µm wide and sometimes formed Y- or H-shaped branches and coils. The coils were ellipsoid, 10.5-23.3 x 36.8-57.4 µm, when observed in a plane view. Extraradical hyphae were (2.7-)3.1(3.4) µm wide and occurred very rarely and in low abundances. In 0.1% trypan blue, arbuscules stained violet white (15A2) to light lilac (16A5), intraradical hyphae violet white (16A2) to pale violet (16A3), coils lilac (16A5) to royal purple (16D8), and extraradical hyphae violet white (16A2) or remained unstained.
 |
 |
 |
 |
 |
 |
 |
 |
 |
In roots of P.lanceolata |
||||||||
PHYLOGENETIC POSITION. Kennedy et al. (1999) concluded that the lack of vesicles and the faintly staining other components of mycorrhizae of Gl. eburneum are untypical of most species of the genus Glomus, but are characteristic for, e. g., Archaeospora trappei (R.N. Ames & Linderman) J.B. Morton & D. Redecker emend. Spain, Appendicispora gerdemannii (S.L. Rose, B.A. Daniels & Trappe) Spain, Oehl & Sieverd. and Diversispora spurca (C.M. Pfeiff., C. Walker & Bloss) C. Walker & Schuessler, of which the former two fungi come from the order Archaeosporales C. Walker & Schuessler and the latter one has recently been accommodated in the family Diversisporaceae C. Walker & Schuessler of the order Diversisporales C. Walker & Schuessler (Schüßler et al. 2001; Spain et al. 2006; Walker and Schüßler 2004). Gamber and Leuchtmann (2007) found the INVAM isolate AZ420A of Gl. eburneum to be in a sister position to Gl. versiforme (P. Karsten) S.M. Berch, being phylogenetically most closely related to D. spurca (Schwarzott et al. 2001).
DISTRIBUTION. In Poland, spores of Gl. eburneum were found in only one trap culture containing a mixture of rhizosphere soil and root fragments taken from under Helichrysum areanarium (L.) Moench growing in maritime sand dunes of the Slowinski National Park (54o45'N, 17o26'E) on 26 June 2003. The occurrence of spores of arbuscular fungi in the field sample was not investigated. The species of arbuscular fungi co-occurring with Gl. eburneum in the trap culture were Archaeospora trappei (R.N. Ames & Linderman) J.B. Morton & D. Redecker emend. Spain and Gl. lamellosum Dalpé, Koske & Tews.
The holotype of Gl. eburneum has been selected from spores extracted from the INVAM culture AZ420A established from a mixture of rhizosphere soil and root fragments of Sporobolus wrightii Monro ex Scribn. growing along the San Pedro River in Arizona, U.S.A. (Kennedy and Morton 1999). Sporobolomyces wrightii is a native grass species found only along rivers and streams of the semiarid regions of south-western North America. Additionally, the same scientists and Stutz et al. (2000) recorded this fungus among roots of other plants growing in other sites of Arizona and Mexico, as well as under different plant species colonizing a dune transect in the Namibia desert.
NOTES. The most distinctive characters of Gl. eburneum are its light-coloured spores filled with dense, opaque oil substance and the semi-flexible, 2-layered spore wall with the outermost layer nonreactive in Melzer's reagent and usually remaining more or less intact in mature spores.
When observed at a low magnification, spores of Gl. eburneum are most reminiscent of those of D. spurca, Gl. albidum C. Walker & L.H. Rhodes, Gl. gibbosum Blaszk., Gl. viscosum Nicol., and Paraglomus occultum (C. Walker) J.B. Morton & D. Redecker. Darker-coloured spores of Gl. eburneum may also be confused with light-pigmented spores of Gl. claroideum N.C. Schenck & G.S. Sm. and Gl. versiforme.
Except for Gl. versiforme, examination of spores crushed in PVLG and PVLG mixed with Melzer's reagent under a compound microscope readily separates Gl. eburneum from all the other species listed above. Considering the spore wall structure, as well as the phenotypic and biochemical properties of its components, the fungus most closely related to Gl. eburneum is D. spurca. Although the opinions of the number of layers overlaying the laminate spore wall layer of D. spurca are contradictory [one layer according to Kennedy et al. 1999 and Morton 2002 vs. two layers as Blaszkowski (2003) stated], the laminate layer and the layer directly overlaying it in the two fungi are almost identical in both their phenotypic and biochemical properties. The main differences between these fungi hide in the persistency of the spore wall layers directly covering the structural laminate layer and the degree of the association of these layers with the laminate layer. Consequently, the second property defines the basic differences in the persistency of the subtending hypha of both fungi.
Although the outer spore wall layer is relatively long-lived and usually retains as a more or less deteriorated structure in mature spores of Gl. eburneum, its spatially corresponding spore wall layer in D. spurca is more persistent and always remains intact in even old spores (Blaszkowski 2003; Kennedy et al. 1999).
In Gl. eburneum, the outer spore wall layer always remains tightly adherent to the laminate spore wall layer, whereas the semi-flexible layer covering the laminate spore wall layer of D. spurca easily separates in crushed spores or balloons when immersed in lactic acid-based mountants (Blaszkowski 2003; Kennedy et al. 1999).
In Gl. eburneum, the structural layer of the subtending hyphal wall (shwl2) continuous with the laminate spore wall layer and being the support of the outer thin wall layer of the subtending hypha gradually thins and expands up to 25 µm below the spore base. In contrast, in D. spurca spores, the laminate spore wall layer abruptly thins and stops to grow at their base and, thereby, it does not create a sufficient support to stabilize the outer subtending hyphal wall layer continuous with the spore wall layer 2 sensu Blaszkowski (2003). Therefore, almost all crushed spores of D. spurca usually lack the subtending hypha, which detaches along with the outer wall layer of these spores.
Finally, in contrast to the opaque, frequently yellow-coloured contents of Gl. eburneum spores, the spore contents of D. spurca consists of transparent oil droplets.
Compared with Gl. albidum described to also form a 2-layered spore wall of similar phenotypic characters (Walker and Rhodes 1981) to those of the wall layers of spores of Gl. eburneum, spores of the latter species do not react in Melzer's reagent (vs. become pink to orange red in G. albidum; Walker and Rhodes 1981).
Two characters of spores of Gl. gibbosum readily separate this fungus from Gl. eburneum. First, while spores of the former species occur in the soil singly, in loose aggregates and conglomerations enclosed by a common hyphal mantle (Blaszkowski 1997, 2003), the latter fungus produces only single spores. Second, the spore wall of Gl. eburneum comprises only two layers, and that of Gl. gibbosum consists of four layers (Blaszkowski 2003). The spore wall of Gl. eburneum lacks the wall layers 2 and 4 of spores of Gl. gibbosum.
Spores of Gl. viscosum are also frequently formed in loose aggregates (Morton 2003; vs. only single spores in Gl. eburneum) and have a more complex wall structure (3-layered) than those of Gl. eburneum (2-layered). The spore wall layer of Gl. viscosum not synthesized by Gl. eburneum is the permanent, semi-flexible, thin layer positioned between a semi-flexible layer forming the spore surface and a laminate layer, both phenotypically similar to the spore wall layers 1 and 2, respectively, of Gl. eburneum.
Of the species compared here, the fungus most diverged morphologically from Gl. eburneum is Par. occultum. These species share only their outermost spore wall layer (Morton 2002; Morton and Redecker 2001). Although these layers in both fungi are of the type of impermanent layers, the longevity of the layer in Gl. eburneum is much higher than in Par. occultum, in which it usually highly deteriorates with age to form a granular structure (Morton 2002), a phenomenon not found in Gl. eburneum. The two other spore wall layers of Par. occultum are uniform and much thinner (both <0.5-1.2 µm thick) than the inner laminate spore wall layer of Gl. eburneum [(2.5-)3.5(-4.4) µm thick].
As mentioned above, darker-coloured spores of Gl. eburneum may also overlap in colour and appearance with light-coloured spores of Gl. claroideum and Gl. versiforme. However, the former two fungi differ fundamentally in the construction of their spore wall, as well as in the phenotypic and biochemical properties of its components. Compared with the simple, 2-layered spore wall of Gl. eburneum, that of Gl. claroideum comprises four layers with the innermost layer staining in Melzer's reagent (Blaszkowski 2003; Morton 2002; Stürmer and Morton 1997; vs. none of the spore wall layers of Gl. eburneum reacts in this reagent). The only layers of the spore wall of Gl. claroideum sharing the phenotypic and biochemical properties of layers 1 and 2 of the Gl. eburneum spore wall are its layers 2 and 3, respectively. The distinctive component of the spore wall of Gl. claroideum is the innermost flexible layer, which is lacking in Gl. eburneum.
Light-coloured spores of Gl. versiforme may be indistinguishable from mature spores of Gl. eburneum when observed under both a dissecting and a compound microscope. Apart from colour, spores of the two species are almost identical in size, as well as in the construction and the phenotypic and biochemical properties of the components of their wall. Moreover, mycorrhizae of both species stain faintly in 0.1% trypan blue (Blaszkowski, pers. observ.; Morton 2002). The only property distinguishing Gl. eburneum and Gl. versiforme is the formation of sporocarps by the latter fungus (Morton 2002).
Apart from the morphological differences characterized above, the species compared here also differ in the phylogenetic position within the phylum Glomeromycota determined based on results of their molecular analyses. For example, Gl. claroideum and Gl. viscosum represent Glomus group B in the family Glomeraceae Piroz. & Dalpé of the order Glomerales J.B. Morton & Benny, and Par. occultum, originally described as Gl. occultum C. Walker, presently is a member of the family Paraglomaceae J.B. Morton & D. Redecker in the Paraglomerales C. Walker & Schuessler (Schüßler et al. 2001). As presented in the section "Phylogenetic position", Gl. eburneum should be a member of the family Diversisporaceae, as Gamber and Leuchtmann (2007) found. Unfortunately, the phylogenetic positions of Gl. albidum and Gl. gibbosum are unknown to date.
REFERENCES
Blaszkowski J. 1997. Glomus gibbosum, a new species from Poland. Mycologia 89, 339-345.
Blaszkowski J. 2003. Arbuscular mycorrhizal fungi (Glomeromycota), Endogone and Complexipes species deposited in the Department of Plant Pathology, University of Agriculture in Szczecin, Poland. http://www.agro.ar.szczecin.pl/~jblaszkowski/.
Gamber H., Leuchtmann A. 2007. Taxon-specific PCR primers to detect two inconspicuous arbuscular mycorrhizal fungi from temperate agricultural grassland. Mycorrhiza 17, 145-152.
Kennedy L. J., Stutz J. C., Morton J. B. 1999. Glomus eburneum and G. luteum, two new species of arbuscular mycorrhizal fungi, with emendation of G. spurcum. Mycologia 91, 1083-1093.
Morton J. B. 2002. International Culture Collection of (Vesicular) Arbuscular Mycorrhizal Fungi. West Virginia University: http://www.invam.caf.wvu.edu/.
Morton J. B., Redecker D. 2001. Two families of Glomales, Archaeosporaceae and Paraglomaceae, with two new genera Archaeospora and Paraglomus , based on concordant molecular and morphological characters. Mycologia 93, 181-195.
Schüßler A., Schwarzott D., Walker C. 2001. A new fungal phylum, the Glomeromycota: phylogeny and evolution. Myc. Res. 105, 1413-1421.
Schwarzott D., Walker C., Schüßler A. 2001. Glomus, the largest genus of the arbuscular mycorrhizal fungi (Glomales) is nonmonophyletic. Mol. Phyl. Evol. 21, 190-197.
Spain J. L., Sieverding E., Oehl F. Revision of the genus Archaeospora and description of Appendicispora gen. novum in the arbuscular mycorrhizae forming Glomeromycetes. Mycotaxon (in press).
Stürmer S. L., Morton J. B. 1997. Developmental patterns defining morphological characters in spores of four species in Glomus. Mycologia 89, 72-81.
Walker C., Rhodes L. H. 1981. Glomus albidus: a new species in the Endogonaceae. Mycotaxon 12, 509-514.
Walker C., Schüßler A. 2004. Nomenclatural clarifications and new taxa in the Glomeromycota. Mycol. Res. 108, 979-982.