Ambispora fennica
C. Walker, Vestberg & Schuessler
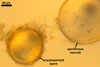
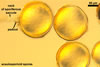
A dimorphic fungus, producing acaulosporioid and glomoid spores. Acaulosporioid spores formed singly, and glomoid ones either singly or in loose aggregates in the soil.
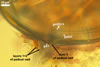
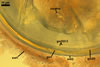
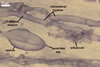
ACAULOSPORIOID SPORES spores origin blastically either at the tip of a short branch (pedicel) of the neck of a sporiferous saccule, a smooth pore, or a slightly raised collar of this neck continuous with a mycorrhizal extraradical hypha. Spores hyaline to deep yellow (4A8); globose to subglobose; (150-)176(-200) µm diam; sometimes ovoid; 150-180 x 175-210 µm.
SUBCELLULAR
STRUCTURE OF ACAULOSPORIOID SPORES consists of a spore wall and two inner germinal walls.
Spore wall composed of three layers
(swl1-3).
|
|
|
|
|
In PVLG |
In PVLG+Melzer's reagent |
Layer 1,
forming the spore surface, evanescent, semipermanent, hyaline to deep yellow (4A8), (5.4-)6.5(-9.1) µm thick, more or less deteriorated in mature spores, often with adherent fine soil particles.
Layer 2 finely laminate, semirigid, hyaline, (2.7-)4.1(-6.1) µm thick.
Layer 3 flexible to semiflexible, hyaline, 0.8-1.5 µm thick, usually tightly adherent to the lower surface of layer 2, and, thereby, difficult to observe.
Germinal wall 1 comprises two fragile, smooth layers (gw1l1 and 2), (0.9-)1.1(-1.4) µm and (1.1-)1.3(-1.7) µm thick, respectively, easily cracking to form line splits or polygonal pieces in crushed spores. Both layers tightly adhere to one another and, hence, usually make an impression that the germinal wall 1 is one-layered.
|
|
|
|
|
|
|
|
|
In PVLG |
In PVLG+Melzer's reagent |
Germinal wall 2 consists of three smooth, hyaline layers
( gw2l1-3).
Layer 1 flexible to semiflexible, (0.7-)1.4(-2.0) µm thick, usually separating from layer 2 in crushed spores.
Layer 2 semiflexible, finely laminate, (3.2-)4.0(-4.4) µm thick.
Layer 3
flexible to semiflexible, ca. 0.5-0.8 µm thick, rarely separating from layer 2 in even vigorously crushed spores.
In Melzer's reagent, only layers 1 and 3 of the spore wall stain red (10A6) to high red (10A8) and greyish yellow (4B4-4C5), respectively.
|
|
|
In PVLG |
In PVLG+Melzer's reagent |
PEDICEL
hyaline to deep yellow (4A8); straight to recurved, cylindrical to funnel-shaped, sometimes slightly constricted; (25.1-)40.7(-50.0) µm long, (14.0-)15.9(-17.4) µm wide at the spore base; positioned 80-130 µm from the base of the saccule; consisting of a hyaline to deep yellow (4A8) wall continuous with the wall of the saccule neck and the spore wall. Pore generally open. In a few spores, the pore was closed by a curved septum continuous with germinal wall 1.
GERMINATION
ORB. Not found.
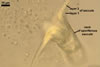
SPORIFEROUS
SACCULE hyaline; globose to subglobose; 199-248 µm diam; collapsing and becoming detached at maturity.
Wall of sporiferous saccule semiflexible, composed of two hyaline layers: an evanescent, 1.2-2.2 µm thick outer layer and a semiflexible, 1.8-3.9 µm thick inner layer.
Saccule neck hyaline; 180-250 µm long, 25-36 µm wide at the base of the saccule, 18-33 µm wide at the spore base, then gradually tapering up to 8-12 µm wide.
CICATRIX. A smooth pore or a slightly raised collar when seen in a cross view; circular; 9-12 µm diam; or ellipsoidal; 4.5-4.9 x 10.0-12.2 µm; when observed in a plane view.
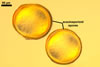
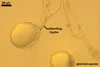
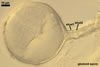
GLOMOID SPORES
spores origin blastically at the tip of hyphae continuous with mycorrhizal extraradical hyphae. Spores hyaline; globose to subglobose; (38-)74(-117) µm diam; sometimes ovoid or irregular; 110-120 x 160-210 µm; with one subtending hypha.
SUBCELLULAR STRUCTURE OF GLOMOID SPORES
consists of a spore wall composed of two hyaline layers (swl1 and 2).
|
|
|
In PVLG |
In PVLG+Melzer's reagent |
Layer 1, forming the spore surface, evanescent, (1.4-)1.6(-1.8) µm thick.
Layer 2
laminate, (2.2-)2.6(-2.9) µm thick, pliable and, thereby, making the spores difficult to crush on microscope slides and observe the details of the structure of their wall.
SUBTENDING HYPHA
hyaline; straight or recurvate; funnel-shaped, rarely cylindrical, occasionally slightly constricted at the spore base, (8.1-)8.9(-9.5) µm wide at the spore base.
Wall of subtending hypha
hyaline; (1.7-)3.4(-5.6) µm thick at the spore base; composed of two layers (shwl1 and 2) continuous with spore wall layers 1 and 2.
Pore open, (3.4-)5.5(-8.1) µm wide.
GERMINATION. Not observed.
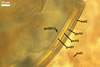
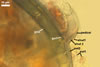
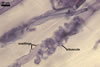
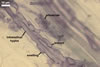
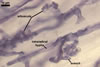
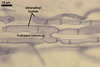
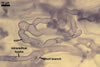
MYCORRHIZAE. In one-species cultures with Plantago lanceolata L. as the host plant, mycorrhizae of Am. fennica consisted of arbuscules, vesicles, as well as intra- and extraradical hyphae. Arbuscules were numerous and evenly distributed along roots. They consisted of short trunks grown from parent hyphae and numerous branches with very fine tips. The numbers of vesicles was variable, from single to numerous, depending on the root fragment examined. They usually were ellipsoid to prolate; 20-45 x 40-85 µm; rarely circular to subcircular; 35-60 µm diam; when observed in a plane view. The top part of the ellipsoid to prolate vesicles was frequently highly constricted and, thereby, resembled the top of a beret. Intraradical hyphae grew along the root axis, were numerous, (4.8-)7.5(-12.5) µm wide, straight or slightly curved, sometimes formed H- or Y-shaped branches, and frequently coils. The straight and curved hyphae were frequently more or less swollen or formed short branches. The coils usually were ellipsoid; 32.5-47.5 x 45.0-80.0 µm; rarely circular; 35.5-47.5 µm diam; when seen in a plane view. Extraradical hyphae were (3.8-)5.8(-9.5) µm wide and occurred in low abundances. In 0.1% trypan blue, arbuscules stained violet white (16A2) to reddish violet (16A6), vesicles pastel violet (16A4) to deep violet (16E8), intraradical hyphae violet white (16A2) to deep violet (16D8), coils pale violet (16A3) to deep violet (16D8), and extraradical hyphae pastel violet (16A4) to greyish violet (16C5).
|
|
|
|
|
|
|
|
|
In roots of P. lanceolata |
PHYLOGENETIC POSITION. Walkers' et al. (2007) phylogenetic analyses showed Am. fennica to belong in a monophyletic clade along with, e. g., Am. gerdemannii (Archaeosporales) and Geosiphon pyriformis (Geosiphonaceae), to which the clade containing Archaeospora trappei represents a sister linage. Archaeospora trappei is the type species of the genus Archaeospora and the family Archaeosporaceae (Morton and Redecker 2001). Another species of the Glomeromycota revealed to belong in the Am. fennica clade is Am. callosa, the former Glomus callosum.
DISTRIBUTION. The holotype of Am. fennica has been selected from spores isolated from a culture with P. lanceolata as the host plant (Walker et al. 2007). This culture was established from an acaulosporioid spore of this fungus extracted from a trap culture with a field soil taken from under pasture grasses and possibly Trifolium pratense L. growing in Kurikka (62º30'N, 22º20'E), Finland. To date, this is the only known site of the occurrence of Am. fennica.
NOTES. The morphological and biochemical properties of spores presented here come from the original description of Am. fennica (Walker et al. 2007), as well as from examination of its spores and fragments of roots obtained from Dr. M. Vestberg, MTT Plant Production Research Laukaa, Vihtavuori, Finland. The material provided was a substrate of one-species culture, named Att550-30, originally established from one acaulosporioid spore of Am. fennica on 13 May 2003.
Examination of morphological and biochemical properties of glomoid spores of Am. fennica made by the author of this book fully confirmed results of Walker's et al. (2007) observations. However, investigations of the same properties of acaulosporioid spores revealed four structures not presented in the original protologue of this species, of which one probably will determine the existence of this species in the future.
First, Walker et al. (2007) overlooked the spore wall layer 3 characterized above. Although this layer rarely separates from the laminate spore wall layer 2, it is slightly thicker that the very fine laminae of this layer, and, hence, more visible. An identical layer has also been revealed in the spore wall of the acaulosporioid morphs of Am. appendicula and Am. gerdemannii (Błaszkowski 2003; Spain et al. 2006). Second, the first germinal wall of spores of Am. fennica is one-layered, as Walker et al. (2007) defined. Examination of this wall under high microscope magnifications rather suggests it to consist of two tightly adherent layers, similarly as in Am. appendicula and Am. gerdemannii (Błaszkowski 2003; Spain et al. 2006). In a few spores, the germinal wall 1 was slightly invaginated at the spore base and the invagination formed a curved septum in the lumen of the pedicel. A similar occlusion of the lumen of a pedicel has been observed in young spores of Am. appendicula and Am. gerdemannii, but not in those of Archaeospora and Acaulospora spp. (Spain et al. 2006). This property distinguishes members of Ambispora from Archaeospora and Acaulospora, the latter two genera also producing acaulosporioid spores (Spain et al. 2006). Third, according to the original description of the fungus discussed here (Walker et al. 2007), its second germinal wall also is one-layered. Meanwhile, analyses of all the spores provided by Dr. M. Vestberg univocally indicated that the discovers of this fungus omitted two layers of this wall, i. e., layers 1 and 3 characterized and illustrated above. Layer 1 is easy to detect, because it is relatively thick (0.7-2.0 µm thick) and almost always more or less separated from the middle layer 2 of this wall in crushed spores. In contrast, layer 3 is very thin (ca. 0.5-0.8 µm thick) and visible as a distinct structure in only vigorously crushed spores. A three-layered structure of similar properties of its components also is the second germinal wall of spores of Am. appendicula and Am. gerdemannii (Błaszkowski 2003; Spain et al. 2006).
In summary, the results of comparative investigations of the morphological and biochemical properties of spores of Am. fennica provided by Dr. M. Vestberg, one of the authors of the paper originally presenting this fungus (Walker et al. 2007), with those collected by the author of this website and those characterized by Spain et al. (2006) suggest that Am. fennica is a synonym of Am. gerdemannii.
Moreover, molecular separateness of the species compared here also is not univocal. Although phylogenetic analyses of sequences from Am. gerdemannii AU215 determined by Wubet et al. (2003 a, b) and Am. fennica revealed by Walker et al. (2007) accommodated these fungi in two subgroups with good support, the Am. gerdemannii sequence (AJ012111) coming from Redecker's et al. (2000) studies separated from them, but with relatively low bootstrap support (Walker et al. 2007).
Thus, to elucidate the doubts presented above, further investigations of morphological, biochemical, and molecular properties of spores of Am. gerdemannii are needed. Such studies should be conducted with specimens propagated in living cultures established from spores of this fungus extracted from the site from which its type was selected.
According to Morton (2002) and Morton and Redecker (2001), the diagnostic properties of mycorrhizae of more ancient members of the Glomeromycota are the lack of vesicles and that their other components stain faintly or do not stain at all in commonly used stains. Meanwhile, Walker et al. (2007) found vesicles in mycorrhizae of both Am. fennica and Am. leptoticha. Almost all root fragments with Am. fennica mycorrhizae stained by the author of this book also contained vesicles. Moreover, all components of these mycorrhizae stained intensively in 0.1% trypan blue. Additionally, vesicles and darkly staining mycorrhizae formed Glomus versiforme and Intraspora schenckii (Błaszkowski 2003), fungi also thought to be within the ancient group of the Glomeromycota (Morton 2002; Sieverding and Oehl 2006). Thus, the taxonomic importance of these properties is doubtful and further research is needed to assess their value, as Walker et al. (2007) concluded.
REFERENCES
Błaszkowski J. 2003. Arbuscular mycorrhizal fungi (Glomeromycota), Endogone, and Complexipes species deposited in the Department of Plant Pathology, University of Agriculture in Szczecin, Poland. http: //www.agro.ar.szczecin.pl/~jblaszkowski/.
Morton J. B. 1993. Problems and solutions for the integration of glomalean taxonomy, systematic biology, and the study of endomycorrhizal phenomena. Mycorrhiza 2, 97-109.
Morton J. B. 2002. International Culture Collection of (Vesicular) Arbuscular Mycorrhizal Fungi. West Virginia University. http://www.invam.caf.wvu.edu/.
Morton J. B., Redecker D. 2001. Two families of Glomales, Archaeosporaceae and Paraglomaceae, with two new genera Archaeospora and Paraglomus, based on concordant molecular and morphological characters. Mycologia 93, 181-195.
Redecker D., Morton J. B., Bruns T. D. 2000. Ancestral lineages of arbuscular mycorrhizal fungi (Glomales). Mol. Phylogenet. Evol. 14, 297-301.
Sieverding E., Oehl F. 2006. Revision of Entrophospora and description of Kuklospora and Intraspora, two new genera in the arbuscular mycorrhizal Glomeromycetes. J. Appl. Bot. Food Qual. 80, 69-81.
Spain J. L., Sieverding E., Oehl F. 2006. Appendicispora: a new genus in the arbuscular mycorrhiza-forming Glomeromycetes, with a discussion of the genus Archaeospora. Mycotaxon 97, 163-182.
Walker C., Vestberg M., Demircik F., Stockinger H., Saito M., Sawaki H., Nishmura I., Schüßler A. 2007. Molecular phylogeny and new taxa in the Archaeosporales (Glomeromycota): Ambispora fennica gen. sp. nov., Ambisporaceae fam. nov., and emendation of Archaeospora and Archaeosporaceae. Mycol. Res. 111, 137-13.
Wubet T., Weiß M., Kottke I., Oberwinkler F. 2003. Morphology and molecular diversity of arbuscular mycorrhizal fungi in wild and cultivated yew (Taxus baccata ). Can. J. Bot. 81, 255-266.
Wubet T., Weiß M., Kottke I., Teketay D., Oberwinkler F. 2003. Molecular diversity of arbuscular mycorrhizal fungi in Prunus africana, an endangered medicinal tree species in dry Afromontane forests of Ethiopia. New Phytol. 161, 517-528.