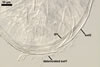

In water
In PVLG
(R.N. Ames & Linderman) J.B. Morton & D. Redecker emend. Spain
|
|
|
|
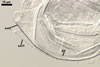
In water |
In PVLG |
||
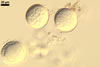
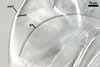
SPORES single in the soil; develop laterally on the neck of a sporiferous saccule; hyaline; globose to subglobose; (37.5-)55.2(-78.0) µm diam.
SUBCELLULAR STRUCTURE OF SPORES consists of a spore wall and one inner germination wall.
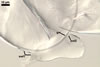 |
 |
 |
 |
 |
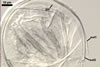 |
In PVLG |
In PVLG+Melzer's reagent |
||||
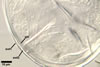
Spore wall composed of two layers (swl1 and 2), frequently wrinkling in intact spores mounted in lactic acid-based mountants.
Layer 1 semiflexible, hyaline, (0.5-)0.6(-0.7) µm thick, usually more or less deteriorated in older spores; this layer is continuous with the wall of the sporiferous saccule.
Layer 2 semiflexible, permanent, hyaline, (0.5-)0.6(-0.7) µm thick.
Germination wall (gw) comprises one semiflexible, hyaline, (1.2-)2.4(-4.2) µm thick layer.
In Melzer’s reagent, neither the spore wall nor any of the two layers of the inner germination wall stain.
GERMINATION ORB. Among the spores of Ar. trappei found by the author of this website, none germinated. According to Spain (2003), Ar. trappei spores germinate by a germ tube developing from an irregular germination structure. This tube emerges through the spore wall.
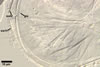
SPORIFEROUS SACCULE hyaline, globose to subglobose; 55-70 µm diam; or ovoid; 50-60 x 70-80 µm; neck 80-120 µm long, 12.5-17.5 µm wide at the saccule, tapering to 5.0-6.5 µm wide at the spore attachment. Saccule usually collapses or falls off in mature spores.
 |
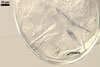 |
In PVLG |
In PVLG+Melzer's |
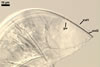
CICATRIX. A scar or sometimes slightly raised collar, when seen in a cross view, circular, 6.0-9.5 µm diam, when observed in a plane view, is formed.
MYCORRHIZAE. In roots of Plantago lanceolata L., mycorrhizae of Ar. trappei consist of arbuscules, as well as intra- and extraradical hyphae staining pale in 0.1% trypan blue.
DISTRIBUTION. In Poland, Ar. trappei has been found in dune soils of the Bledowska Desert (50o22’N, 19o34’E; Błaszkowski et al. 1999; Błaszkowski et al. 2002a), the Slowinski National Park (54o45’N, 17o26’E; Tadych and Błaszkowski 2000a), the Vistula Bar (54o24'N, 19o30'E; Błaszkowski et al. 2002b), and in soils of the Tuchola Forests (53o46’N, 17o42’E-53o40’N, 17o54’E; Tadych and Błaszkowski 2000b).
Archaeospora trappei has originally been known from Lolium longiflorum Thunb. fields located along southern Oregon and northern California coastal areas (Ames and Linderman 1976). This fungus has a worldwide distribution and is adapted to a wide range of host plants and soil conditions. It has been found in, e. g., Australia, Brazil, Cuba, Japan, Namibia, South Africa, Scotland (Morton and Redecker 2001), many regions of the U.S.A. (Bever et al. 1996; Hetrick and Bloom 1983; Morton and Redecker 2001; Schenck and Kinloch 1980; Schenck and Smith 1981; Walker et al. 1982), Iceland (Greipsson et al. 2002), Germany, Switzerland and France (Blaschke 1991; Oehl et al. 2005), United Kingdom (Merryweather and Fitter 1998), Israel (Błaszkowski et al. 2001; Haas and Menge 1990), China (Gai et al. 2006), and Central Taiwan (Wu and Chen 1986).
NOTES. Spores of Ar. trappei most resemble those of Intraspora schenckii (Sieverd. & S. Toro) Oehl & Sieverd. Both species form hyaline spores of an identical subcellular structure. However, spores of Ar. trappei form laterally on the neck of a sporiferous saccule, and those of I. schenckii develop inside the neck (Sieverding and Oehl 2006; Sieverding and Toro 1987).
Similarly as members of the genus Appendicispora Spain, Oehl & Sieverd., Ar. trappei is a dimorphic fungus producing acaulosporioid and glomoid spores (Sieverding and Oehl 2006; Spain et al. 2006). Its acaulosporioid morph described above forms spores identically to those of species of the gnus Acaulospora. According to Spain (2003), glomoid spores of Ar. trappei form blastically and intercalary, are hyaline, 22.5-31.25 µm diam, and their wall consists of two layers, each 1-1.5 µm thick. Inoculation of roots of Zea mays L. with these spores failed.
In the literature, two opinions of the number of walls in spores of Ar. trappei exist. All the spores examined by the author of this website contained only a 3-layered spore wall, similarly as spores characterized by Morton and Redecker (2001). In contrast, Spain (2003) observed in germinating spores of this species a 2-layered spore wall and four 1-layered inner walls, of which the innermost one formed upon germination. The author of this website did not find any germinating spore of this fungus. Therefore, further studies are needed to explain this inconsistency.
Archaeospora trappei is the only member of the genus Archaeospora. Two other species originally included in this genus (Morton and Redecker 2001), i. e., Ar. appendicula (Sain, Sieverd. & N.C. Schenck) [erroneously considered to be Ar. leptoticha (N.C. Schenck & S.M. Sm.) J.B. Morton & D. Redecker by Morton and Redecker (2001) as Spain et al. (2006) concluded] and Ar. gerdemannii S.L. Rose, B.A. Daniels & Trappe have been transferred to the newly established genus Appendicispora and renamed Ap. appendicula (Spain, Sieverd. & N.C. Schenck) Spain, Oehl & Sieverd. and Ap. gerdemannii (S.L. Rose, B.A. Daniels & Trappe) Spain, Oehl & Sieverd., respectively (Spain et al. 2006).
The main arguments used to separate the two species mentioned above from the original composition of the genus Archaeospora were (1) different production of their acaulosporioid spores (in Appendicispora spp., they form on a pedicel (appendix) branched from the neck of a sporiferous saccule vs. Ar. trappei produces sessile spores, directly on the neck of a sporiferous saccule), (2) differences in the subcellular structure of spores of members of these genera; germinating spores of Ap. appendicula consist of a 3-layered spore wall and two 2-layered inner germination walls (Spain et al. 2006), whereas germinating spores of Ar. trappei comprise a 2-layered spore wall and four 1-layered inner walls, and (3) inconsistency in the mode of germination of spores of these fungi; spores of Ap. appendicula, the only species in which this process has so far been recognized, germinate through a germ tube emerging through the lumen of their pedicel, and the germ tube of germinating spores of Ar. trappei emerges through their spore wall.
REFERENCES
Ames R. N., Linderman R. G. 1976. Acaulospora trappei sp. nov. Mycotaxon 3, 565-569.
Bever J., Morton J. B., Antonovics J., Schultz P. A. 1996. Host-dependent sporulation and species diversity of arbuscular mycorrhizal fungi in a mown grassland. J. Ecol. 84, 71-82.
Blaschke H. 1991. Multiple mycorrhizal associations of individual calcicole host plants in the alpine grass-heath zone. Mycorrhiza 1, 31-34.
Błaszkowski J., Tadych M., Madej T., Adamska I., Czerniawska B., Iwaniuk A. 1999. Acaulospora mellea and A. trappei, fungi new to the Mycota of Poland. Acta Mycol. 34, 41-50.
Błaszkowski J., Tadych M., Madej T. 2002a. Arbuscular mycorrhizal fungi (Glomales, Zygomycota) of the Bledowska Desert, Poland. Acta Soc. Bot. Pol. 71, 71-85.
Błaszkowski J., Adamska I., Czerniawska B. 2002b. Arbuscular mycorrhizal fungi (Glomeromycota) of the Vistula Bar. Acta Mycol. 37, 39-62.
Błaszkowski J., Tadych M., Madej T., Adamska I., Iwaniuk A. 2001. Arbuscular mycorrhizal fungi (Glomales, Zygomycota) of Israeli soils. Mat. II Polsko-Izraelskiej Konf. Nauk. nt. „Gospodarowanie zasobami wodnymi i nawadnianie roslin uprawnych”. Przeglad naukowy Wydz. Inz. Ksztalt. Srod. 22, 8-27.
Gai J. P., Christie P., Feng G., Li X. L. 2006. Twenty years of research on biodiversity and distribution of arbuscular mycorrhizal fungi in China: a review. Mycorrhiza 16, 229-239.
Haas J. H., Menge J. A. 1990. VA-mycorrhizal fungi and soil characteristics in avocado (Persea americana Mill.) orchard soils. Plant and Soil 127, 207-212.
Hetrick B. A. D., Bloom J. 1983. Vesicular-arbuscular mycorrhizal fungi associated with native tall grass prairie and cultivated winter wheat. Can. J. Bot. 61, 2140-2146.
Merryweather J., Fitter A. 1998. The arbuscular mycorrhizal fungi of Hyacinthoides non-scripta. I. Diversity of fungal taxa. New Phytol. 138, 117-129.
Morton J. B., Redecker D. 2001. Two new families of Glomales, Archaeosporaceae and Paraglomaceae, with two new genera Archaeospora and Paraglomus, based on concordant molecular and morphological characters. Mycologia 93, 181-195.
Oehl F., Sieverding E., Ineichen K., Ris E.-A., Boller T., Wiemken A. 2005. Community structure of arbuscular mycorrhizal fungi at different soil depths in extensively and intensively managed agroecosystems. New Phytol. 165, 273-283.
Schenck N. C., Kinloch R. A. 1980. Incidence of mycorrhizal ungi on six field crops in monoculture on a newly cleared woodland site. Mycologia 72, 445-456.
Schenck N. C., Smith G. S. 1981. Distribution and occurrence of vesicular-arbuscular mycorrhizal fungi on Florida agricultural crops. Soil and Crop Sci. Soc. Florida 40, 171-175.
Sieverding E., Oehl F. 2006. Revision of Entrophospora and description of Kuklospora and Intraspora, two new genera in the arbuscular mycorrhizal Glomeromycetes. J. Appl. Bot. Food Qual. 80, 69-81.
Sieverding E., Toro S. T. 19887. Entrophospora schenckii: a new species in the Endogonaceae from Colombia. Mycotaxon 28, 209-214.
Spain J. L. 2003. Emendation of Archaeospora and its type species, Archaeospora trappei. Mycotaxon 87, 109-112.
Spain J. L., Sieverding E., Oehl F. 2006. Appendicispora: a new genus in the arbuscular mycorrhiza-forming Glomeromycetes, with a discussion of the genus Archaeospora. Mycotaxon 97, 163-182.
Tadych M., Błaszkowski J. 2000a. Arbuscular fungi and mycorrhizae (Glomales) of the Slowinski National Park, Poland. Mycotaxon 74, 463-483.
Tadych M., Błaszkowski J. 2000b. Arbuscular mycorrhizal fungi of the Brda river valley in the Tuchola Forests. Acta Mycol. 35, 3-23.
Walker C., Mize C. W., Mcnabb, Jr. H. S. 1982. Populations of endogonaceous fungi at two locations in central Iowa. Can. J. Bot. 60, 2518-2529.
Wu Ci-G., Chen Z.-Ch. 1986. The Endogonaceae of Taiwan. I. A preliminary investigation on Endogonaceae of bamboo vegetation at Chi-Tou areas, Central Taiwan. Taiwania 31, 65-88.