
Oehl, Sykorova & Sieverd.
|
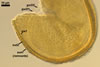
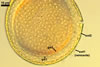
SPORES single in the soil; origin laterally on the neck of a sporiferous saccule; pale yellow (3A3) to deep yellow (4A8; Blaszkowski, pers. observ.) to orange-brown (Oehl et al. 2006); globose to subglobose; (53-)65-85(-97) µm diam; rarely ovoid to irregular; (53-)60-81(-91) x 62-87(-110) µm.
SUBCELLULAR STRUCTURE OF SPORES consists of a spore wall and two inner germination walls.
 |
 |
 |
 |
 |
 |
 |
 |
 |
| In PVLG |
||||||||
 |
 |
 |
 |
 |
In PVLG+Melzer's reagent |
||||
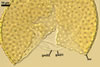
Spore wall composed of three layers (swl1-3).
Layer 1, forming the spore surface, evanescent, hyaline, 0.5-1.0 µm thick, usually highly deteriorated or completely sloughed in mature spores.
Layer 2 laminate, pale yellow (3A3) to deep yellow (4A8; Blaszkowski, pers. observ.) to orange-brown (Oehl et al. 2006); (1.7)2.7(-3.9) µm thick, ornamented with evenly distributed, circular, truncated conical pits, (1.5-)1.9(-2.8) µm diam when observed in a plane view, (0.7-)1.6(-2.2) µm deep when seen in a cross-sectional view, spaced (2.2-)3.6(-4.4) µm apart.
Layer 3 semiflexible, concolorous with layer 2, 0.5-1.3 µm thick, usually tightly adherent to the lower surface of layer 2 and, hence, difficult to detect, especially when thinner than 1 µm.
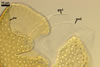
Germination wall 1 composed of two semiflexible, hyaline layers, of a total thickness of 0.5-1.2 µm; these layers rarely separate from one another and, thereby, are exceptionally difficult to observe.
Germination wall 2 comprises three adherent hyaline layers (gw2l1-3).
Layer 1 flexible, smooth, ca. 0.5 µm thick.
Layer 2 flexible, 1.2-2.0 µm thick.
Layer 3 flexible, ca. 0.5 µm thick, tightly adherent to the inner surface of layer 2 and always very difficult to detect.
In Melzer's reagent, only layer 2 of the germination wall 2 stains reddish white (7A2) to pastel red (7A4). However, this staining reaction is not visible in all specimens.
GERMINATION ORB. Not found.
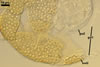
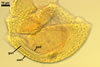
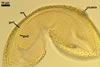
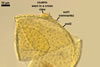
SPORIFEROUS SACCULE consists of a saccule and a neck.
Saccule hyaline; globose to subglobose; ca. 65-80 µm diam; or ellipsoid; 65-75 x 75-92 µm; formed at a distance of 40-80 µm from the spore.
Neck tapering from the saccule towards its end.
Saccule wall composed of one hyaline layer, 1.0-2.1 µm thick. The saccule usually collapses after the spore has formed and is usually detached from mature spores isolated from the soil.
 |
 |
In PVLG |
|
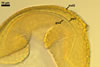
CICATRIX. A slightly raised tapering collar, circular, (5.0-)7.0-12.0 µm diam when observed in a plane view, extending into the lumen of the neck of the sporiferous saccule up to 0.8-2.2(-3.0) µm when seen in a cross view. In developing spores, the collar surrounds a pore that becomes closed by some of the laminae of the spore wall layer 2 with time. While the surface of the closed pore is smooth, the outer surface of the wall of the collar is pitted similarly as the upper surface of the spore wall layer 2.
MYCORRHIZAE. In the field, A. alpina has been associated with plant communities of alpine grasslands dominated by Carex curvula All., C. ferruginea Scop., Nardus stricta L., Sesleria caerulea (L.) Scop., and Trisetum flavescens (L.) P. Beauv. Attempts to grow this fungus in one-species pot cultures failed. Thus, mycorrhizae of A. alpina remain unknown.
PHYLOGENETIC POSITION. Phylogenetic analyses placed A. alpina in a clade not related with clades comprising other species of the genus Acaulospora characterized molecularly to date (Oehl et al. 2006). The most unexpected result of these analyses was that A. alpina was not similar molecularly to A. paulinae, a species forming pitted spores of morphological characters most resembling those of spores of the former fungus, and A. lacunosa, another fungus producing pitted spores of a similar pigmentation that may also relatively easily be confused with spores of A. alpina.
DISTRIBUTION. Spores of A. alpina have occurred abundantly in soils of a pH of 3.5-6.5 of different grasslands of the Swiss Alps dominated by the plant species listed in the section Mycorrhizae and located 1800 to 2700 m above see level (Oehl et al. 2006). This fungus has been more rarely found in sites located at 1300-1800 m above see level and has not so far been revealed in the Alps below 1300 m and in the lowlands of Switzerland.
In Poland, ......
Switzerland and Poland are the only countries where A. alpina was found to date.
NOTES. Except for ...... spores of A. alpina (microphotographs .....; slides .......) isolated from ....., the other spores showed here come from Dr. F. Oehl's, Institute of Botany, University of Basel, Switzerland, collection (slides no. .......).
The description and illustrations of A. alpina were prepared based on the original protologue (Oehl et al. 2006) and results of examination of spores of this fungus found in Poland and those loaned by Dr. F. Oehl.
The properties of spores of A. alpina described by Oehl et al. (2006) generally agreed with those determined by me. Small differences were found only in the thickness of the spore wall layer 2 [(1.7-)2.7(-3.9) µm thick vs. 2.0-3.0 µm thick in Oehl et al. 2006] and the distance between the pits of this layer [(2.2-)3.6(-4.4) µm apart vs. (3.0-)4-6(-7) µm apart in Oehl et al. 2006]. Additionally, spores of this fungus coming from both Poland and Switzerland were slightly paler [pale yellow (3A3) to deep yellow (4A8)] than those originally characterized (dark yellow, orange to brown; Oehl et al. 2006). However, all the specimens of this fungus examined were mounted in either PVLG or its mixture with Melzer's reagent, both containing lactic acid that always lightens coloured spore components of arbuscular fungi (Blaszkowski, pers. observ.; Morton 1986b).
Additionally, except for the properties of layer 2 of the germination wall 2, those of some other components of the subcellular structure of A. alpina spores were given here only based on the original description of this fungus (Oehl et al. 2006), because the available specimens lacked them or they were invisible.
First, none of the spores examined was associated with its sporiferous saccule. Second, the spore wall layer 3 and layer 3 of the germination wall 2 were not visible at all. Second, the germination wall 1 in all spores seemed to be one-layered, and not composed of two components. However, one of the microphotographs of Oehl et al. (2006) clearly shows two layers in this wall. Many freshly extracted spores usually need to be crushed to stratify such a wall (Blaszkowski, pers. observ.).
In the original protologue (Oehl et al. 2006), the properties of layer 2 of the germination wall 2 presented regard only its thickness and reactivity in Melzer's reagent. Unfortunately, there is no information of the phenotype of this layer, a character having a high taxonomic importance in recognition of members of the Glomeromycota (Franke and Morton 1994). In all the spores investigated by me, layer 2 of the germination wall 2 seemed to be flexible, and not plastic, as in A. paulinae, A. lacunosa, and many other Acaulospora spp. Although the plasticity of this layer is visible best in crushed spores freshly extracted from living cultures and the visibility of this character usually diminishes in spores preserved in lactic acid-based mountants, it generally is recognizable. Moreover, Morton (1986b) concluded that a plastic layer distinguishes that it always stains intensively in Melzer's reagent, except for changes induced by mountants or spore preservation. Thus, the faint or no staining of this layer in Melzer's reagent in A. alpina indicates it to be not plastic.
The distinctive properties of A. alpina are its small, darkly coloured spores ornamented with evenly distributed circular pits when seen in a plane view and the second germination wall consisting of three layers, of which the outermost one is smooth and the middle one either stains faintly or does not stain at all in Melzer's reagent.
Of the other species of the genus Acaulospora forming pitted spores, the fungi most similar to A. alpina are A. paulinae, A. undulata, and A. taiwania because of their small spores. However, spores of A. paulinae and A. undulata generally are paler in colour [hyaline to pale yellow (3A3) and hyaline to subhyaline, respectively; Blaszkowski 2003; Blaszkowski, pers. observ.] than those of A. alpina [pale yellow (3A3) to deep yellow (4A8) to orange-brown; Blaszkowski, pers. observ; Oehl et al. 2006; Sieverding 1988]. Although spores of A. taiwania also resemble in colour those of the fungus discussed here, they are formed in sporocarps (Hu 1988) rather than singly in the soil as in A. alpina.
Another species producing pitted and yellow (4A6) to deep yellow (4A8) spores is A. lacunosa, but its globose spores may be much larger (up to 140 µm diam vs. up to 100 µm diam in A. alpina; Blaszkowski 2003; Blaszkowski, pers. observ.; Morton 1986a; Oehl et al. 2006).
The most distinct differences between the species listed above reside in the phenotypic and biochemical properties of the components of the subcellular structure of their spores. While the laminate layer of the spore wall of A. alpina is ornamented with evenly distributed truncated conical pits of a diameter of 1.5-2.5 µm when observed in a plane view (Blaszkowski, pers. observ.; Oehl et al. 2006), that of A. paulinae is ornamented with concave depressions that are larger (2.0-2.5 x 3.0-3.5 µm) and less regularly distributed in this layer than in the former species (Blaszkowski 2003).
Circular to irregular and relatively larger depressions also ornament the laminate spore wall layer of A. undulata (Blaszkowski, pers. observ.; Sieverding 1988).
Other important taxonomically differences between some of the species listed above regard the properties of layers 1 and 2 of the second germination wall of their spores. Layer 1 of the second germination wall of A. alpina spores is smooth, and layer 2 of this wall stains faintly or does not stain at all in Melzer's reagent (Blaszkowski, pers. observ.; Oehl et al. 2006). In contrast, layer 1 of the germination wall 2 of spores of A. paulinae and A. lacunosa is covered with granular excrescences (a beaded layer), and its layer 2 reacts intensively in this reagent (Blaszkowski 1988; Morton 1986a, 2002).
Acaulospora undulata differentiates only one germination wall of properties partly fitting those of the germination wall of Archaeospora trappei (Blaszkowski, pers. observ.; Oehl et al. 2006), now the only member of the genus Archaeospora (Spain et al. 2006). However, the former fungus does not form the germinal wall corresponding with the first germinal wall of all the species listed above. This and the faint staining mycorrhizae of A. undulata suggest that this fungus A. undulata should be transferred to the genus Archaeospora, as Oehl et al. (2006) concluded.
The other known members of the genus Acaulospora with pitted spores are A. cavernata, A. foveata, A. excavata, and A. scrobiculata. Compared with spores of A. alpina, those of A. scrobiculata generally are lighter coloured [yellowish white (3A2) to pale yellow (3A3) vs. pale yellow (3A3) to deep yellow (4A8) to orange-brown; Blaszkowski, pers. observ.; Oehl et al. 2006] and larger [(70-)88(-100) µm diam when globose vs. (90-)120(-135) µm diam or (80-)120(-160) µm diam; Blaszkowski 2003; Blaszkowski, pers. observ; Morton 2002], and those of the other species are larger and darker in colour: yolk yellow (4B4) to light brown (6D8) and (110-)140(-170) µm diam when globose in A. cavernata, pale ochraceous to ochre to orange and 115-200 x 100-165 µm in A. excavata, and red-orange to dark red-brown and (240-)289(-360) µm diam in A. foveata; Blaszkowski 2003; Morton 2002; Ingleby et al 1994.
Except for A. alpina and A. cavernata, the outermost layer of the germination wall 2 of the other species considered is of the type of a beaded layer. In A. alpina, its upper surface is smooth, whereas in A. cavernata it is ornamented with knobby swellings resembling an inner covering of postal letters (Blaszkowski 2003; Blaszkowski, pers. observ.; Oehl et al. 2006).
Finally, the middle layer of the second germination wall of spores of A. alpina stains faintly or does not stain at all in Melzer's reagent (Blaszkowski, pers. observ.; Oehl et al. 2006). In contrast, a layer corresponding with this layer in A. cavernata, A. excavata, A. foveata, and A. scrobiculata always stains intensively in this reagent (Blaszkowski 2003; Ingleby et al. 1994; Morton 2002).
REFERENCES
Blaszkowski J. 1988. Three new vesicular-arbuscular mycorrhizal fungi (Endogonaceae) from Poland. Bull. Pol. Ac. Sci. Biol. Sci. 36, 10-12.
Blaszkowski J. 2003. Arbuscular mycorrhizal fungi (Glomeromycota), Endogone, and Complexipes species deposited in the Department of Plant Pathology, University of Agriculture in Szczecin, Poland.http://www.agro.ar.szczecin.pl/~jblaszkowski/.
Franke M., Morton J. B. 1994. Ontogenetic comparisons of arbuscular mycorrhizal fungi Scutellospora heterogama and Scutellospora pellucida: revision of taxonomic character concepts, species descriptions, and phylogenetic hypotheses. Can. J. Bot. 72, 122-134.
Hu H. T. 1988. Study on the endomycorrhizae of China fir (Cunninghamia lanceolata Hooker) and taiwania (Taiwania cryptomerioides). Q. J. Chin. For. 21, 45-72.
Ingleby K., Walker C., Mason P. A. 1994. Acaulospora excavata sp. nov. - an endomycorrhizal fungus from Cote D'Ivoire. Mycotaxon 50, 99-105.
Morton J. B. 1986. Three new species of Acaulospora (Endogonaceae) from high aluminum, low pH soils in West Virginia. Mycologia 78, 641-648.
Morton J. B. 1986. Effects of mountants and fixatives on wall structure and Melzer's reaction in spores of two Acaulospora species (Endogonaceae). Mycologia 78, 787-794.
Morton J. B. 2002. International Culture Collection of Arbuscular and Vesicular-Arbuscular Mycorrhizal Fungi. West Virginia University: http://www.invam.caf.wvu.edu/.
Oehl F., Sýkorová Z., Redecker D., Wiemken A. 2006. Acaulospora alpina, a new arbuscular mycorrhizal fungal species characteristic for high mountainous and alpine regions of the Swiss Alps. Mycologia 98, 286-294.
Spain J. L., Sieverding E., Oehl F. 2006. Appendicispora: a new genus in the arbuscular mycorrhiza-forming Glomeromycetes, with a discussion of the genus Archaeospora. Mycotaxon 97, 163-182.
Sieverding E. 1988. Two new species of vesicular arbuscular mycorrhizal fungi in the Endogonaceae from tropical high lands of Africa. Angew. Bot. 62, 373-380.