6-week
old
|
|
6-week
old |
|
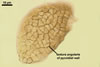
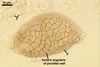
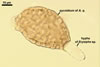
COLONIES slow-growing, golden yellow (5B7) to raw umber (5F8), 12-23 mm diam, with an irregular margin and superficial or immersed, at first hyaline, later dark orange (5B8) to yellowish brown (5D8), branched, septate aerial mycelium when grown on potato dextrose agar (PDA) at a room temperature and 6-week old.
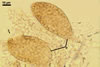
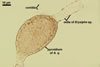
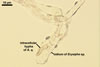
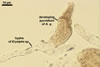
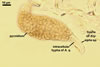
PYCNIDIA (p) superficial, in/on or around hyphae and young cleistothecia of species of the order Erysiphales, separate, globose, elongated to pyriform, 40-90(-105) x (10-)20-50(-65) µm, pale yellow (4A3) to golden yellow (5B8) when young, golden yellow (5B7) to yellowish brown (6E8) at maturity, unilocular, with a thick wall.
 |
 |
 |
 |
 |
 |
 |
 |
 |
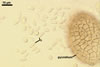
Wall of pale brown textura angularis, with no distinct ostiolum; dehiscence by apical rupture of pycnidium.
Conidiophores absent.
Conidiogenous cells enteroblastic, phialidic, discrete, smooth, hyaline, 4.5-5.5 µm wide, formed directly from the pycnidial wall cells.
 |
 |
 |
 |
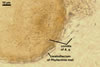
CONIDIA (c) hyaline, widely ellipsoidal to cylindrical, (4.5-)6.8(-10.5) x (3.0-)4.1(-6.0) µm.
 |
 |
 |
HOST AND DISTRIBUTION. Ampelomyces quisqualis has been found associated in ca. 65 fungal species of eight genera of the order Erysiphales (powdery mildew fungi) in different parts of the world (Kiss 1998). In Poland, A. quisqualis has been revealed on 18 species of the genus Erysiphe, two of Microsphaera, eight of Sphaerotheca, two of Podosphaera, and one each of Oidium, Phyllactinia, and Uncinula (Czerniawska 2001; Czerniawska unpubl. data; Madej and Antoszczyszyn 1965). Phyllactinia mali (Duby) U. Braun was a host of A. quisqualis earlier not given in the literature.
NOTES. Although descriptions of over 40 species of the genus Ampelomyces exist in the literature (Sutton 1980), the only species commonly accepted was A. quisqualis (Kiss 1998). However, recent molecular studies indicated the existence of more than one Ampelomyces species in the nature (Kiss 1997; Kiss and Nakasone 1998). According to Kiss and Nakasone (1998), Ampelomyces spp. have their phylogenetic position in the order Dothideales (Ascomycota). When Ampelomyces internal transcribed spacer sequences were analyzed by cladistic methods with the sequences of other ascomycetous fungi, they formed two lineages in the Dothideales. Slow-growing Ampelomyces isolates formed a clade with Leptosphaeria microscopica P. Karsten and L. nodorum E. Müller, whereas fast-growing Ampelomyces isolates formed a clade with Epicoccum purpurascens Ehrenb. ex Schlecht., a fungus known to have a pycnidial, Phoma-like state (Arenal et al. 2000). Consequently, Kiss et al. (2004) suggested that the fast-growing isolates were, in fact, Phoma species, whereas the true Ampelomyces isolates were always slow-growing in culture and always produced intracellular pycnidia in powdery mildew mycelia.
Currently, A. quisqualis is considered a common pycnidial intracellular mycoparasite of probably all powdery mildews world-wide. However, according to Kiss (1996), the extremely low incidence of A. quisqualis in powdery mildews of plant species of the family Poaceae and its not uniform occurrence in different geographical regions suggest this mycoparasite to play no significant role in natural control of grass and cereal mildews.
 |
 |
 |
 |
 |
 |
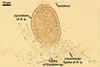 |
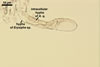
Developmental stages of
pycnidia of Ampelomyces quisqualis |
||||||
Conidia of A. quisqualis are produced in pycnidia developed intracellularly in the mycelia of powdery mildew fungi. The conidia are dispersed within the plant canopy by rain-splash or water run off from plant surfaces. For long distances, A. quisqualis can also spread as hyphal fragments in parasitized and detached powdery mildew conidia. When these parasitized air-borne conidia land close to any powdery mildew colony under humid conditions, the outgrowing hyphae of A. quisqualis can penetrate their mycelia. After penetration, the hyphae of A. quisqualis invade the host mycelia internally and produce pycnidia, mainly in the conidiophores and young ascocarps of powdery mildews. Sometimes, A. quisqualis also produces pycnidia in the invaded hyphal cells.
 |
 |
 |
In contrast to some other fungal hyperparasites, e. g., Trichoderma spp. producing antifungal compounds, acting as mycoparasites, inducing plant defense mechanisms, and stimulating plant growth (Howell 2003), A. quisqualis invades and kills the parasitized powdery mildew cells by causing a rapid degeneration of the cytoplasm (Hashioka and Nakai 1980). The early stage of mycoparasitism is biotrophic, but the invaded cytoplasm then dies and a necrotrophic interaction begins. No production of any toxins has been recognized (Kiss 2003). Phillipp et al. (1984) found that the parasitized powdery mildew colonies can continue their growth, but their sporulation stopped soon after A. quisqualis penetrated their mycelia.
 |
 |
Ampelomyces quisqualis overwinters in the form of pycnidia produced saprotrophically in the senescent or dead plant tissues, in the parasitized ascocarps of powdery mildew fungi, and in the mycelium of powdery mildews produced during the previous season (Kiss et al. 2004).
Data on the efficacy of A. quisqualis as a biocontrol agent are contradictory (Kiss et al. 2004). To maintain control, the growth and spread of the powdery mildew fungal hosts must be slower than that of A. quisqualis. Ampelomyces quisqualis destroys its powdery mildew hosts slowly (during 5-7 days) compared with other microbial antagonists, e. g., species of Trichoderma and Verticillium. Therefore, when the sporulation and spread of powdery mildews are intense, A. quisqualis can not control them completely. Nevertheless, this mycoparasite now is one of the most advanced in commercial development of a fungal biocontrol agent for plant pathology.
REFERENCES
Arenal F., Platas G. Monte E., Pelaez F. 2000. ITS sequencing support for Epicoccum nigrum and Phoma epicoccina being the same biological species. Myc. Res. 104, 301-303.
Czerniawska B. 2001. Studies on the biology and occurrence of Ampelomyces quisqualis in the Drawski Landscape Park (NW Poland). Acta Mycol. 36, 191-201.
Hashioka Y., Nakai Y. 1980. Ultrastructure of pycnidial development and mycoparasitism of Ampelomyces quisqualis parasitic on Erysiphales. Trasn. Mycol. Soc. Japan 21, 329-338.
Howell C. R. 2003. Mechanisms employed by Trichoderma spp. in the biological control of plant diseases: the history and evolution of current concepts. Plant Disease 87, 4-10.
Kiss L. 1996. Graminicolous powdery mildew fungi as new natural hosts of Ampelomyces mycoparasites. Can. J. Bot. 75, 680-683.
Kiss L. 1997. Genetic diversity in Ampelomyecs isolates, hyperparasites of powdery mildew fungi, inferred from RFLP analysis of the rDNA ITS region. Mycol. Res. 101, 1073-1080.
Kiss L. 1998. Natural occurrence of Ampelomyces intracellular mycoparasites in mycelia of powdery mildew fungi. New Phytol. 140, 709-714.
Kiss L. 2003. A review of fungal antagonists of powdery mildews and their potential as biocontrol agents. Pest Manag. 59, 475-483.
Kiss L., Nakasone K. K. 1998. Ribosomal DNA internal transcribed spacer sequences do not support the species status of Ampelomyces quisqualis, a hyperparasite of powdery mildew fungi. Curr. Genet. 33, 362-367.
Kiss L., Russel J. C., Szentiványi O., Xu X., Jeffries P. 2004. Biology and biocontrol potential of Ampelomyces mycoparasites, natural antagonists of powdery mildew fungi. Biocontrol Sci. Technology 14, 635-651.
Madej T., Antoszczyszyn S. 1965. Ameplomyces quisqualis Ces. (Cicinobolus cesatii De Bary) w Szczecinie. Biul. Inst. Ochr. Rosl. 30, 65-76.
Phillipp W. D., Grauer U., Grossmann F. 1984. Ergänzende Untersuchungen zur biologischen und integrierten Bekämpfung von Gurkenmehltau under Glass durch Ampelomyces quisqualis. Z. Pflkrankh. Pflschutz 91, 438-443.
Sutton B. 1980. The Coelomycetes. Fungi imperfecti with pycnidia, acervuli, and stromata. Commonwealth Mycological Institute. Kew, Surrey, England.