_jpg.jpg)
-r_jpg.jpg)


6-day
old
10-day old
|
|
|
|
6-day
old |
10-day old |
||
COLONIES fast-growing, white (5A1), low, flaky, covering the whole area of potato dextrose agar (PDA) of a diameter of 10 cm when 6-day old, becoming grey (5B1) to brownish grey (5C2) and forming sclerotia at the border of the PDA medium when 10-day old and grown at a room temperature.
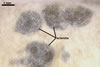
CONIDIOPHORES (cp) produced from mycelium or sclerotia, usually formed in tufts, 750 µm to over 2 mm long, smooth-walled, grey-brown to brown, 18-23 µm wide in the lower part, hyaline to pale brown and branched in the upper part; each branch ended with a hemispherical or spherical swelling, 5-12 µm diam, having minute sterigmata (s), i. e., spore-bearing projections.
 |
 |
 |
 |
 |
 |
 |
 |
 |
 |
 |
 |
 |
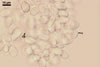 |
CONIDIA (c) of two types: macro- and microconidia.
Macroconidia globose, ellipsoidal, or egg-shaped, smooth, hyaline to pale brown, usually with a protuberant hilum, 8-14 x 6-9 µm.
Microconidia not observed.
 |
 |
SCLEROTIA shield-like, lenticular to irregular, brown to black, 0.2-0.5(15) mm diam, consisting of densely packed medulla and a pseudoparenchymatous, dark brown to black cortical layer of cells, 5-10 µm diam.
SUBSTRATE AND DISTRIBUTION. Botrytis cinerea is a facultative parasite commonly inhabiting plant debris and occasionally attacking a very wide range of living plant species, especially those of the families Asteraceae and Liliaceae (Domsch et al. 1980). This fungus has frequently been found among microorganisms colonizing seeds, leaves, and inflorescences of different plant species (Blaszkowski 1994a, b, c, 1995; Blaszkowski, Piech 2002; Domsch et al. 1980; Lacicowa 1963; Neergaard 1977; Piech, Blaszkowski 1993).
Botrytis cinerea has a worldwide distribution (Domsch et al. 1980). However, it prefers humid temperate and subtropical regions.
NOTES. The teleomorph of B. cinerea is Botryotinia fuckeliana (De Bary) Whetz. (Holliday 1989).
The genus Botryotinia Whetzel has been separated from the genus Sclerotinia Fuckel by having a more compact, gelatinized, sclerotial medulla and a Botrytis anamorph (Smith et al. 1988).
 |
 |
 |
Botrytis cinerea causes the so-called "grey mold" of different cultivated plant species. This disease is one of the most commonly occurring plant diseases in the world. The plants most frequently attacked by B. cinerea are, e. g., strawberry (Fragaria x ananassa Duchesne), tomato (Lycopersicon esculentum Mill.), bean (Phaseolus vulgaris L.), rose (Rosa spp.), cyclamen (Cyclamen purpurascens Mill.), chrysanthemum (Chrysanthemum spp.), dahlia (Dahlia spp.), raspberry (Rubus idaeus L.), grape-vine (Vitis vinifera L.), currants (Ribes spp.), sunflower (Helianthus annuus L.), and apples (Malus domestica Borkh.; Agrios 1988).
The disease symptoms caused by B. cinerea appear on all above ground plant parts and in all plant developmental stages.
 |
 |
Botrytis cinerea overwinters in the forms of mycelium growing on decaying plant debris and sclerotia in the soil or on the soil surface. Seed infection does occur, but seems to be unimportant (Smith et al. 1988).
The conditions favouring the growth of B. cinerea, its sporulation, spore release, germination, and establishment of infection are 18-24oC and damp weather. This fungus remains its activity even if the temperatures are between 0 and 10oC and, thereby, causes considerable losses on crops stored for long periods.
Although B. cinerea may penetrate directly into its plant host, the germ tubes of germinated conidia usually enter into plant tissues through wounds.
Apothecia with ascospores of the teleomorph B. fuckeliana are infrequently produced from sclerotia.
REFERENCES
Agrios G. N. 1988. Plant pathology, 3rd edition, Academic Press, INC. San Diego, New York, Berkeley, Boston, London, Sydney, Tokyo, Toronto.
Blaszkowski J. 1994. The occurrence of Septoria nodorum Berk. and associated mycoflora in seeds of wheat cultivated in the Szczecin voivodeship. Acta Mycol. 29, 43-52.
Blaszkowski J. 1994. The influence of foliar fungicides on the mycoflora of seeds of Triticum aestivum. Acta Mycol. 29, 141-145.
Blaszkowski J. 1994. The influence of fungicides on the mycoflora of leaves of Triticum aestivum L. Acta Mycol. 29, 147-157.
Blaszkowski J. 1995. Effects of foliar fungicides on the mycoflora of glumes of Triticum aestivum. Acta Mycol. 30, 41-48.
Blaszkowski J., Piech M. 2002. Comparison of seed-borne fungal communities of naked and husked oats and barley. Phytopath. Pol. 24, 71-74.
Brooks F. T. 1953. Plant diseases. Geoffrey Cumberlege. Oxford University Press. London, new York, Toronto.
Domsch K. H., Gams W., Anderson T. 1980. Compendium of soil fungi. Acad. Press. London-New York-Toronto-Sydney-San Francisco.
Fassatiová O. 1983. Grzyby mikroskopowe w mikrobiologii technicznej. Wydawnictwo Naukowo-Techniczne. Warszawa.
Holliday P. 1989. A dictionary of plant pathology. Cambridge University Press. Cambridge, New York, New Rochelle, Melbourne, Sydney.
Lacicowa B. 1963. Badania nad morfologia i biologia Fusarium poae (Pk.) Wr. oraz patogenicznoscia tego gatunku wzgledem siewek pszenicy. Ann. Univ. Mariae Curie-Sklodowska 18, 419-439.
Neergaard P. 1977. Seed pathology. The Macmillan Press Ltd. London.
Piech M., Blaszkowski J. 1993. The influence of the degree of reproduction on the mycoflora of winter XTriticosecale seeds. Bull. Pol. Ac. Sci. Biol. Sci. 41, 393-399.
Smith I. M., Dunez J., Lelliott R. A., Phillips D. H., Archer S. A. 1988. European handbook of plant diseases. Blackwell Scientific Publications.