







LEAF SPOTS on the upper leaf side, at first single, small, circular to subcircular, yellow brown to purple, with radiating strands of mycelium below the cuticule, later numerous, confluent, covering a large leaf area, more uniform and dark brown to black, with highly rugged borders and small, circular to prolate, blisters arranged either at random, when distant away from the leaf border, or in rings, when located at its border. The leaf tissue around the spots turns yellow and frequently, when severely infected, entire leaves become yellow and fall off prematurely.
|
|
|
|
|
|
|
|
 |
 |
 |
 |
 |
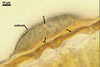
ACERVULI (a) form between the outer wall and cuticule of the epidermis, consist of a basal wall and of conidiogenous cells lining the upper surface of the wall. When young, the acervuli are completely covered with the cuticule. The developing conidiogenous cells and conidia push up the cuticule and, thereby, produce blisters on the upper leaf side. Later, the blisters burst and expose the conidiomata.
Wall (w) composed of thick-walled, tightly adherent angular cells.
Conidiogenous cells (cc) short, cylindrical or conical, hyaline.
 |
 |
 |
CONIDIA (c) hyaline, clavate, 1-septate, constricted at the septum, 10-24 x 4.5-7 µm. The upper cell is frequently slightly narrowed and sharpened at the top, whereas the bottom cell is usually widened and rounded at the base.
PLANT HOST AND DISTRIBUTION. Marssonina rosae attacks different plant species of the genus Rosa.
The fungus occurs wherever both cultivated and wild roses grow (Brooks 1953).
NOTES. The teleomorph of M. rosae is Diplocarpon rosae Wolf. Diplocarpon rosae has not been found in Poland to date.
Marssonina rosae is the causal agent of black spot of roses (Agrios 1988; Brooks 1953; Kochman 1973, 1986). Although M. rosae mainly affects leaves, the fungus can also occasionally attack young stems and flower stalks (Brooks 1953).
In Poland, M. rosae overwinters as mycelium and conidia in infected leaves and canes (Kochman 1973). The fungus can also persists saprotrophically on fallen leaves, where conidia are formed in subepidermal acervuli (Smith et al. 1988). The primary infections of leaves in the spring are produced by conidia, whose germ tubes penetrate into the leaves directly. Under favourable conditions, i. e., 15-25oC, conidia may germinate within 9 hours. Conidia are formed throughout the growing season and cause repeated infections during warm and wet weather (Agrios 1988). They are dispersed by rain splash and possibly by insects (Smith et al. 1988).
REFERENCES
Agrios G. N. 1988. Plant pathology, 3rd edition, Academic Press, INC. San Diego, New York, Berkeley, Boston, London, Sydney, Tokyo, Toronto.
Brooks F. T. 1953. Plant diseases. Geoffrey Cumberlege. Oxford University Press. London, New York, Toronto.
Kochman J. 1973. Fitopatologia. PWRiL. Warszawa.
Kochman J. 1986. Zarys mikologii dla fitopatologów. Wydawnictwo SGGW-AR. Warszawa.
Smith I. M., Dunez J., Lelliott R. A., Phillips D. H., Archer S. A. 1988. European handbook of plant diseases. Blackwell Scientific Publications.