GERMINATION.
Not
observed.
MYCORRHIZAE.
In the field, spores of Gl. multiforum have been found associated
with vesicular-arbuscular mycorrhizal roots of Ammophila
arenaria (L.) Link, Plantago
major L., and Poa trivialis L. (Blaszkowski and Tadych 1997;
Blaszkowski, pers. observ.).
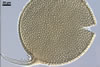
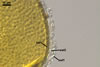
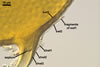
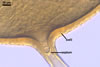
Glomus multiforum
formed vesicular-arbuscular mycorrhizae in pot cultures with P. lanceolata
L. and Sorghum sudanense (Staph.) Piper. Arbuscules were numerous
and evenly distributed along the root fragments examined. Intraradical hyphae occurred abundantly and frequently formed coils. All the structures stained
intensively in 0.1% trypan blue.
|
 |
|
|
|
|
|
In roots
of P. lanceolata
|
DISTRIBUTION.
In
Poland, Gl. multiforum has been found only in one soil-root sample
collected under a mixture of P. major and P. trivialis growing
in a meadow located in Izdebki (53o17'N, 17o19'E; Blaszkowski
and Tadych 1997).
Additionally,
one of the authors of this website, J. Blaszkowski, found this fungus in the root zone of A. arenaria
growing in dunes of the Mediterranean Sea located ca. 15 km from Adana
(36o43N, 34o59'E), Turkey.
NOTES.
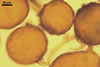
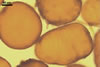
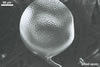
The unique feature of Gl. multiforum spores is their
ornamentation consisting of evenly distributed pits in wall layer 3.
Immature spores are
hyaline and have a wall composed of two, thin layers. Because these layers
tightly adhere to each other, they are frequently seen as a one-layered structure.
At times, layer 1 thickens and gradually deteriorates. Layer 2 thickens and
produces evenly distributed ingrowths. It is rigid and resembles a unit wall
sensu Walker (1983). In the next stage of spore development, layer
1 further decomposes and begins to slough, although almost all spores possess
this layer. Layer 2 remains unchanged. The laminated layer is gradually synthesized
from thin, coloured laminae, of which some outer ones form pits due to the
tight adherence to the ingrowths of layer 2. In the next stage, layer 1 may
be completely sloughed. Layer 2 is more or less decomposed. In many spores,
layers 1 and 2 resemble a structure composed of many layers, of which the
innermost one does not stain in Melzer’s reagent. Layers 1 and 2 of
older spores may by completely sloughed, although most spores coming from
both the field and pot cultures possess layer 2.
Differentiation of spore
wall layers in Gl. multiforum proceeded similarly as in Gl. caledonium
(Nicol. & Gerd.) Trappe & Gerd. (Morton 1996). The innermost laminated
layer 3 of this new fungus began to form when the outer layers 1 and 2 completed
their phenotypic properties. The best evidence of such a sequence in the development
of these spore wall layers are the pits of layer 3, which form based on the
ornamentation of spore wall layer 2. The presence in one-species cultures
of Gl. multiforum of both spores with very shallow pits and those
without them suggests that formation of layer 3 proceeds after a pause. During
this period layer 2 probably undergoes decomposition that causes its ingrowths
to become too soft to impress clear or any pits in the laminated layer 3.
When seen under a dissecting
microscope, spores of Gl. multiforum most closely resemble those
of Gl. geosporum (Nicol. &
Gerd.) Walker, Gl. mosseae
(Nicol. & Gerd.) Gerd. & Trappe and Gl.
verruculosum Blaszk. (Blaszkowski, pers. observ.; Blaszkowski and
Tadych 1997; Nicolson and Gerdemann 1968; Morton 2000). The three species
form spores similar in size, shape, colour, and have a relatively broad subtending
hypha occluded by a curved septum. However, Gl. mosseae spores sometimes
occur in sporocarps with a peridium, whereas those of Gl. geosporum,
Gl. multiforum, and Gl. verruculosum are ectocarpic. Additionally,
most mature spores of Gl. multiforum usually are darker than those
of Gl. mosseae.
Examination of spores
under a light microscope readily separates Gl. multiforum from Gl.
geosporum, Gl. mosseae, and Gl. verruculosum, as well
as from all the other known species of the genus Glomus. Only Gl.
multiforum produces pitted spores. Smooth spores of Gl. multiforum
may be distinguished from those of Gl. geosporum and Gl. verruculosum
with no ornamentation on the inner surface of spore wall layer 3 based on
their subcellular wall structure and reaction in Melzer's reagent. Although
the spore wall structure of all these species consists of three layers, spores
of both Gl. geosporum and Gl. verruculosum lack the middle
semiflexible wall layer of Gl. multiforum spores. On the other hand,
spores of Gl. multiforum do not form the innermost semirigid
wall layer of spores of Gl. geosporum and Gl. verruculosum.
The outermost spore wall layer of Gl. geosporum and Gl. multiforum
stains in Melzer's reagent, whereas that of Gl. verruculosum spores
is nonreactive in this reagent. Smooth spores of Gl. multiforum and
Gl. mosseae may be indistinguishable due to the similarity in appearance
and spore wall structure with the outermost mucilaginous layer staining in
Melzer’s reagent. However, the spore wall of the former fungus usually
is much thicker (6.6-14.2 µm) than that of the latter species (2-7 µm;
Gerdemann and Trappe 1974; Blaszkowski, pers. observ.).
Spores of Gl. multiforum
examined under a light microscope are most similar to those of Archaeospora
leptoticha (Schenck & Nicol.) Morton & Redecker. The
latter fungus produces spores with a Glomus-like pedicel and has
the exact same ornamentation pattern on wall layers 2 and 3 as does the former
species (Morton and Redecker 2001). However, Ar. leptoticha forms
spores laterally on the neck of a sporiferous saccule, whereas those of Gl.
multiforum develop terminally by swelling hyphal tip. The sporiferous
saccule of fungi of the genus Archaeospora frequently becomes completely
detached (Morton 2000), and hence their spores, especially those of Ar.
leptoticha with its persistent pedicel, may be confused with spores of
Glomus spp. having a fragile subtending hypha, as, e. g., in Gl.
albidum Walker & Rhodes (Walker and Rhodes 1981; Blaszkowski, pers.
observ.). However, the subtending hypha of Gl. multiforum
is much longer than that of Ar. leptoticha. Additionally, compared
with Gl. multiforum spores possessing a spore wall comprising three
layers: two sloughing layers and a laminated layer, those of Ar. leptoticha
have a 4-layered spore wall: an outer wall composed of a sloughing layer adherent
to two unit layers and a separable inner flexible, coriaceous layer (coriaceous
wall sensu Walker 1983; Morton 2000; Morton and Redecker 2001).
Other arbuscular fungi
forming pitted spores similar in size, shape and colour to those of Gl.
multiforum are Acaulospora
cavernata Blaszk., Ac. excavata Ingley & Walker, and
Ac. foveata Trappe & Janos (Blaszkowski 1989; Janos and Trappe
1982; Ingleby et al. 1994). The presence of the persistent funnel-shaped subtending
hypha in Gl. multiforum (vs. sessile spores in Ac. cavernata,
Ac. excavata, and Ac. foveata) and inner walls in Ac.
cavernata, Ac. excavata, and Ac. foveata (vs. no inner
wall in Gl. multiforum) easily distinguishes the four fungi.
The high morphological
variability of Gl. multiforum spores may cause difficulties in the
recognition and, thereby, determination of the occurrence of this fungus,
especially based on spores recovered from the field.
REFERENCES
Blaszkowski J. 1989.
Acaulospora cavernata (Endogonaceae) - a new species from Poland
with pitted spores. Crypt. Bot. 1, 204-207.
Blaszkowski J., Tadych
M. 1997. Glomus multiforum and G. verruculosum, two new
species from Poland. Mycologia 89, 804-811.
Gerdemann J. W., Trappe
J. M. 1974. The Endogonaceae in the Pacific Northwest. Myc. Memoir 5, 1-76.
Ingleby K., Walker C., Mason P. A. 1994. Acaulospora excavata sp.
nov. - an endomycorrhizal fungus from Cote D'Ivoire. Mycotaxon 50, 99-105.
Janos D. P., Trappe
J. M. 1982. Two new Acaulospora species from tropical America. Mycotaxon
15, 515-522.
Morton J. M. 1996. Redescription
of Glomus caledonium based on correspondence of spore morphological
characters in type specimens and a living reference culture. Mycorrhiza 6,
161-166.
Morton J. B. 2000. International
Culture Collection of Arbuscular and Vesicular-Arbuscular Mycorrhizal Fungi.
West Virginia University.
Morton J. B., Redecker
D. 2001. Two families of Glomales, Archaeosporaceae and Paraglomaceae, with
two new genera Archaeospora and Paraglomus, based on concordant
molecular and morphological characters. Mycologia 93, 181-195.
Nicolson T. H., Gerdemann
J. W. 1968. Mycorrhizal Endogone species. Mycologia 60, 313-325.
Walker C. 1983. Taxonomic
concepts in the Endogonaceae: spore wall characteristics in species descriptions.
Mycotaxon 18, 443-455.
Walker C., Rhodes L.
H. 1981. Glomus albidus: a new species in the Endogonaceae. Mycotaxon
12, 509-514.